Invasion History
First Non-native North American Tidal Record: 1916First Non-native West Coast Tidal Record: 1967
First Non-native East/Gulf Coast Tidal Record: 1916
General Invasion History:
The bryozoan Victorella pavida was first identified from the London docks by Saville-Kent (1870). It has subsequently been recorded from many European ports (Carrada and Sacchi 1964; D'Hondt 1983; Occhipinti Ambrogi and d'Hondt 1981; Partaly 1979), the Black and Caspian Seas (Mordukhai-Boltovskoi 1964), Israel (Massard et al. 1992), India, Thailand (Wood et al. 2006), Japan, California (Menon and Nair 1967; Winston 1977; Carlton 1979), and the Atlantic, Gulf and West Coasts of North America (Osburn 1944; Everitt 1975; Ruiz et al. unpublished data).
Carlton (1979), while acknowledging taxonomic difficulties, has suggested that V. pavida is a species of Indo-Pacific origin which has been transported worldwide by ships, first to Britain and elsewhere in Europe, then to the east coast of North and South America, and later to the West Coast (Lake Merritt, Oakland, CA; 1967) by way of Japan. However, Mordukhai-Boltovskoi (1964) regarded this bryozoan as native to the Ponto-Caspian region and introduced worldwide, as was the hydroid Cordylophora caspia. It appears that careful comparisons of morphology of live organisms and genetic studies will be needed to determine the origins of this species or species complex. Victorella pavida is probably introduced in England and northern Europe. Marine biology was such a widespread interest there in the late 1800's, it is likely that native populations would have been recognized much earlier if they existed. We treat this bryozoan as introduced in Northern Europe and along the Pacific coast of North America where it has a scattered distribution, and is largely confined to ports. We consider it introduced to the Mediterranean and Ponto-Caspian regions, the Western Atlantic, and Eastern Pacific, and probably native to the Indo-Pacific region, where it occurs both in fresh and brackish waters (Wood et al. 2006).
North American Invasion History:
Invasion History on the West Coast:
On the Pacific Coast of North America, Victorella pavida is a definite introduction. It was found in Lake Merritt, Oakland, California, in 1967 (Carlton 1979). It had not been collected in other San Francisco Bay locations until it was found in the Turning Basin of the Petaluma River in 2004 (Cohen et al. 2005). Victorella pavida has also been found in the Tijuana River, San Diego, and in Elkhorn Slough (de Rivera et al. 2005).
Invasion History on the East Coast:
We consider Victorella pavida to be inteoduced on the East and Gulf coasts, despite some difficulty m its identification. Some of the V. pavida identified from the Atlantic coast of the US, including specimens illustrated by Osburn (1944) from Chesapeake Bay, may be Tanganella appendiculata, presumably a native North American species, first described in 1982 from Martha's Vineyard, Massachusetts. Victorella pavida and Tanganella spp. can only be distinguished in living material, and consequently many 'Victorella' identifications are uncertain (Jebram and Everitt 1982). We will refer to records which could be V. pavida, T. appendiculata, or other related species as 'V. pavida (s.l.)' (for 'sensu lato', meaning in the broad sense). Winston (personal communication) examined live 'V. pavida (s.l.)' specimens from fouling plates from Baltimore Harbor, but has not yet determined the identity of the organisms.
On the East Coast, Victorella pavida was first reported from the lower Chesapeake Bay, off Cape Charles City, Virginia, in a biological survey of the Bay in 1915-1922 (Cowles 1930; Osburn 1932). However, this bryozoan may have been present earlier. The first collections of Victorella pavida were in the mid-to-lower Bay, at locations between James Island, MD and Cape Charles City, VA (Cowles 1930; Osburn 1932). Osburn (1944) later reported that it ranged over 3-27 ppt, with an optimum around 10-14 ppt. He also noted that 'Where it is abundant, it is a nuisance in fouling the bottoms of boats. Oysters and barnacles may be covered to the edges of their shells' (Osburn 1944). Bibbins (1893), studying Cordylophora in Furnace Creek, Baltimore Harbor, noted 'dense and beautiful growths of bryozoans', most likely Victorella pavida (s. l.). since this is the only bryozoan occurring there now (Ruiz et al., unpublished). Victorella pavida (s. l.) has also been reported from North Carolina (in 1973, Dean and Bellis 1975), South Carolina (Calder and Maturo 1978); and the Indian River Lagoon, Florida (in 1975, Winston 1982). Chesapeake Bay appears to be its northern limit on the Atlantic Coast.
Invasion History on the Gulf Coast:
The first published record of Victorella pavida in the Gulf of Mexico region was in the New Orleans area on the Lake Pontchartrain Causeway, at the south end of Lake Pontchartrain (1971, Poirrier and Mulino 1977). Other Gulf records were from Madisonville, Louisiana on the Tchefuncte River (1974, Poirrier and Mulino 1977), from Lake Borgne (Everitt 1973); and from Davis Bayou, in Ocean Springs, Mississippi (1975, Poirrier and Mulino 1977). It has also been collected in Texas, in Galveston Bay, Corpus Christi Bay (2003, Ruiz et al., unpublished data).
Invasion History Elsewhere in the World:
Victorella pavida is apparently introduced in northern Europe. It was described from the London docks on the Thames River (type locality, Saville-Kent 1870; Carrada and Sacchi 1969). It was subsequently reported from many other European ports, including Ostend, Belgium, the Zuiderzee, Netherlands, in Bergues, Dunkerque, and Caene in Normandy, France, and the Aber estuary, in Roscoff (Brittany), France (Carrada and Sacchi 1969). Victorella pavida is no longer found at some locations, including the Thames estuary, and the Zuiderzee, Netherlands (Carrada and Sacchi 1969). Currently, in the British Isles, Victorella pavida is only found in Swanpool: a brackish water lagoon near Falmouth in Cornwall (MarLin 2009; Carter et al. 2010). In the Baltic, V. pavida was first found in the Greifswald Bodden, a lagoon of the Baltic in Germany, in 1880 (Nehring 2001). It has been found in several locations in Germany, Sweden, Poland, and Finland (Carrada and Sacchi 1969).
Victorella pavida (s.l) is widely reported in warmer parts of the world, where we regard it as cryptogenic. It is recorded from estuaries in Mondevideo and Rio de la Plata, Uruguay and Sao Paulo and Rio de Janeiro, Brazil (Marcus 1955). It is also known from coastal lagoons on the Atlantic coast of Morocco, and throughout the Mediterranean (Carrada and Sacchi 1969), the Black and Caspian Seas (Mordukhai-Boltovskoi 1964; Carrada and Sacchi 1969), the east and west coasts of India (Menon and Nair 1967; Rao and Ganapati 1975), fresh and brackish waters of Thailand (Wood et al. 2006), and Matsushima Bay, Japan (Toriumi 1944, cited by Winston 1977). Some of these records may consist of closely related species and genera.
Description
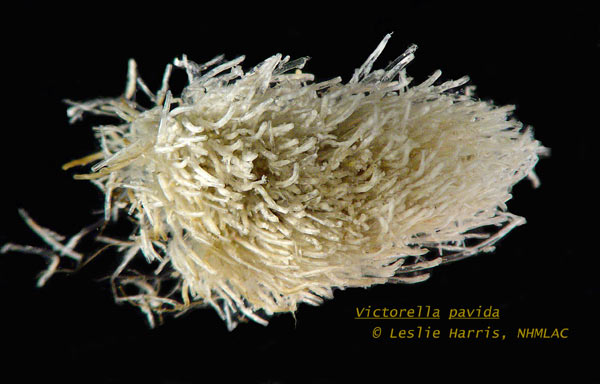
Victorella pavida is an erect, un-calcified bryozoan, found in fresh and brackish waters. Its colonies form diffuse branching chains, or dense clumps of tubular zooids, which are closely or loosely attached to a substratum. Colonies can form a dense mat 3-6 mm high as creeping colonies, but can grow into branching forms 15 mm high, and can spread over a radius of 100-150 mm. The zooids are variable in shape and size. On bare surfaces, it forms creeping colonies of zooids 300-430 µm long and about 150-160 µm thick. Buds form from the basal portion of the zooids. About 180-220 mm of the length, in the distal portion, is the peristomial tube- when the polypide retracts the tube contains only the setae, which form a collar around the mouth and tentacles. As the colony becomes more crowded, the zooids become erect and longer, reaching 1000-1200 µm, joined by short proximal tubes. In erect zooids, budding can occur from the midpoint of the zooid. The new branching zooid can have a long proximal tube, up to 1500 µm, leading to a wider portion, about 450-500 µm, where the polypide is housed. The polypide has a collar of setae and eight tentacles around the mouth. Two of the tentacles are bent sharply away from the others. The pharynx and esophagus leads to the expanded cardia region of the stomach- this species lacks a gizzard. Sexually reproductive zooids develop an intratentacular organ for releasing eggs. The embryos develop in an invagination of the dorsal wall of the vestibule (description from: Osburn 1944; Carrada and Sacchi 1964; Ryland 1965; Winston 1982; Hayward 1985; Wood et al. 2006; Winston and Hayward 2012). Under stressful conditions, colonies form modified zooids, called hibernacula, which are brown, dome-shaped structures, 160-201 µm in diameter, which serve as propagules for surviving cold, hypoxia, or other stresses (Carter et al. 2010).
Taxonomy - There is considerable uncertainty in the taxonomy and identification of Victorellidae and related bryozoans. Many important morphological features cannot be seen in preserved material and the colonies can change form as they grow (Jebram and Everitt 1982). Victorella pavida has been perceived either as a naturally cosmopolitan species (Hyman 1959), or as one widely introduced worldwide (Carlton 1979), but some populations originally identified as V. pavida may be other species of Victorella or other genera (Tanganella, Nolella, Bulbella, etc.) (Jebram and Everitt 1982).
In the genus Victorella, the cardial sphincter is located at the midpoint of the cardial region (rather than at the beginning of the digestive caecum, as in Tanganella) (Jebram and Everitt 1982). However, gut morphology is difficult to observe, and its taxonomic value is untested (Wood et al. 2006). In the genus Bulbella, only erect zooids occur, and the larvae are brooded externally. Based on other criteria, at least six species of Victorella have been recognized, but their validity is uncertain (Wood et al. 2006). Osburn's (1932) identification of Victorella pavida from Chesapeake Bay was somewhat provisional; comments from British Museum taxonomists Harmer and Kirkpatrick were: 'Your example differs a good deal from the typical material; but on the other hand resembles closely the figures of Kraepelin. We are both inclined to think that you would not be far wrong if you named your specimen 'Victorella pavida.' So the determination stands and must remain until better preserved material can be properly studied'. Osburn (1944), Poirrer and Mulino (1977b) and other authors had noticed a correlation between decreasing salinity and decreased branching of 'Victorella pavida' colonies. Jebram and Everitt (1982) suggest that this may represent differences among cryptic species. Wood et al. (2006) observed a high degree of variability in cultured and wild colonies in freshwater in Thailand, depending on crowding, and possibly seasonal and environmental influences.
The possible distinctions among multiple species potentially confused with Victorella pavida, are likely to be missed in earlier work and in most routine fouling surveys. Given the uncertainty over species identification, records from the Chesapeake and other localities, as well as life history and community data, should be regarded as 'V. pavida 'sensu lato' (in the broad sense, ignoring possible cryptic speciation- 's. l.' for short) unless we have noted them as more closely identified. Dr. Judy Winston has identified 'Victorella pavida s. l.', resembling Osborn's material from Chesapeake Bay, including material from the Smithsonian Environmental Research Center 'Invasions Group' settling plates (Winston personal communication).
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Bryozoa | |
| Class: | Gymnolaemata | |
| Order: | Ctenostomata | |
| Suborder: | Carnosa | |
| Superfamily: | Paludicelloidea | |
| Family: | Victorellidae | |
| Genus: | Victorella | |
| Species: | pavida |
Synonyms
Potentially Misidentified Species
Bowebankia spp. bud from the stolons, and have a gizzard. All 8 tentacles are uniformly straight or curved.
Bowerbankia imbricata
None
Bowerbankia teritia
None
Bulbella abscondita
This species was described from Martha's Vineyard, Massachusetts (Jebram and Everitt 1982).
Tanganella appendiculata
This species was described from Martha's Vineyard, Massachusetts (Jebram and Everitt 1982)
Tanganella muelleri
This species has been reported from European waters (Baltic-Mediterranean). There is one report from Cape Cod, Massachusetts.
Victorella pseudoarachnida
This species was described from Salton Sea, California. The distinctive feature is the occurrence of branched cystid appendages, at the junctions between zooids (Jebram and Everitt 1982). This species has not been reported from any other location. It is possible that this feature, occurring occasionally in other species (Jebram and Everitt 1982) may be influenced by the unique environment of the Salton Sea,
Ecology
General:
Life History- Victorella pavida is a soft-bodied bryozoan, which forms creeping or bush-like colonies composed of many individual zooids. The zooids feed by extending the ciliated tentacles of the lophophore as a funnel, creating a current, and driving food particles into their mouths. The food is guided along the tentacles and through the pharynx by the cilia. Larger food particles can be moved or captured by flicking or contracting the tentacles. The zooids are hermaphroditic, and produce large yolky eggs, which hatch into lecithotrophic larvae that are planktonic for short periods (less than 1-2 days). Larvae settle on a substrate and metamorphose into the first zooid of a colony, an ancestrula (Hyman 1959; Barnes 1983). Under stressful conditions, including falling temperatures, and hypoxia, zooids form structures called hibernacula, which survive when the colony dies back. Hibernacula remain attached to the substratum, and are not viable for more than a year (unlike the statoblasts of most freshwater bryozoans) (Carter et al. 2010).
Ecology- Victorella pavida has been found attached to vegetation, woody debris, jetties, shells, pilings, and boats (Osburn 1944; Maturo 1957). This bryozoan has been reported from freshwater in India, Thailand, and Louisiana, but is best known from brackish waters (Osburn 1944; Carrada and Sacchi 1964; Everitt 1975; Poirrier, and Mulino 1977; Massard et al. 1992; Wood et al. 2006). Populations in different regions seem to vary greatly in salinity tolerance, which could reflect either intraspecific genetic differences or cryptic speciation. Colonies in England grew poorly at salinities below 9 PSU (Carter et al. 2010), while in Thailand this bryozoan is widespread in fresh water (Wood et al. 2006). In India, colonies that settled during the monsoon period at 0.3-5.2 PSU, had 100% 24-hour survival, while those that settled in the post-monsoon period at 14.6-22.4 PSU did not survive in freshwater (Menon and Nair 1967).
Food:
Phytoplankton, detritus
Trophic Status:
Suspension Feeder
SusFedHabitats
| General Habitat | Grass Bed | None |
| General Habitat | Coarse Woody Debris | None |
| General Habitat | Oyster Reef | None |
| General Habitat | Marinas & Docks | None |
| General Habitat | Rocky | None |
| General Habitat | Canals | None |
| General Habitat | Vessel Hull | None |
| Salinity Range | Oligohaline | 0.5-5 PSU |
| Salinity Range | Mesohaline | 5-18 PSU |
| Salinity Range | Polyhaline | 18-30 PSU |
| Salinity Range | Limnetic | 0-0.5 PSU |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Epibenthic | None |
Life History
Tolerances and Life History Parameters
| Minimum Temperature (ºC) | 0 | Field data- Nauman and Cory 1968 |
| Maximum Temperature (ºC) | 37.8 | Field data (Thermal effluent)- Nauman and Cory 1969) |
| Minimum Salinity (‰) | 0 | Field Data- Carrada and Sacchi 1964; Cowles 1930; Osburn 1944; Menon and Nair 1967; Everitt 1975; Poirrier and Mulino 1977b; Winston 1977; Wood et al. 2006 |
| Maximum Salinity (‰) | 36 | Experimental data (Carter et al. 2010); 22-30 PSU, Field Data- Carrada and Sacchi 1964; Cowles 1930; Osburn 1944; Menon and Nair 1967; Everitt 1975; Poirrier and Mulino 1977b; Winston 1977; Wood et al. 2006). |
| Minimum Duration | 0.1 | Larval period- Carrada and Sacchi 1964; Hyman 1959 |
| Maximum Duration | 1 | Larval period- Carrada and Sacchi 1964; Hyman 1959 |
| Maximum Height (mm) | 15 | Osburn 1944 |
| Broad Temperature Range | None | Cold temperate-Tropical |
| Broad Salinity Range | None | Nontidal limnetic-Polyhaline |
General Impacts
Economic Impacts
Industry - Victorella pavida (s. l.) is an important fouling organism on power plants and other industrial equipment in estuarine waters of the Patuxent River, Chesapeake Bay (Nauman and Cory 1969) and the Sea of Azov (Partaly 1979).
Boating - Victorella pavida (s. l.) is 'a nuisance fouling species on the bottom of boats' in Chesapeake Bay (Osburn 1944; Lippson and Lippson 1984).
Ecological Impacts
Competition - Victorella pavida (s. l.) is presumed to compete for space in mesohaline fouling communities with the introduced hydroids Cordylophora caspia and Garveia franciscana, which are also frequently dominant in fouling communities (Cory 1967; Nauman and Cory 1969; Abbe 1987; Ruiz et al. unpublished data).
Habitat Change - Victorella pavida (s. l.) colonies provide habitat for a wide variety of small motile animals (amphipods, polychaetes, etc.) in Chesapeake Bay (Ruiz et al. unpublished observations).
Regional Impacts
| NA-ET3 | Cape Cod to Cape Hatteras | Ecological Impact | Competition | ||
| Victorella pavida (s. l.) is one of the dominant fouling organisms in summer in Chesapeake Bay (Osburn 1944; Cory 1967; Lippson and Lippson 1984; Abbe 1987) and is presumed to compete with other fouling taxa for space in mesohaline fouling communities, including native barnacles and the introduced hydroids Cordylophora caspia and introduced Garveia franciscana, which are also frequently dominant in fouling communities (Cory 1967; Nauman and Cory 1969; Abbe 1987; Ruiz et al. unpublished data). |
|||||
| NA-ET3 | Cape Cod to Cape Hatteras | Ecological Impact | Habitat Change | ||
| Victorella pavida (s. l.) colonies provide habitat for a wide variety of small motile animals (amphipods, polychaetes, etc.) in Chesapeake Bay (Ruiz et al. unpublished observations). | |||||
| NA-ET3 | Cape Cod to Cape Hatteras | Economic Impact | Shipping/Boating | ||
| Boating - Victorella pavida (s. l.) is 'a nuisance fouling the bottom of boats' in Chesapeake Bay (Osburn 1944; Lippson and Lippson 1984). |
|||||
| NA-ET3 | Cape Cod to Cape Hatteras | Economic Impact | Industry | ||
| Victorella pavida (s. l.) is an important fouling organism in power plants and other industrial users of estuarine waters in the Patuxent River, Chesapeake Bay (Nauman and Cory 1969). | |||||
| MED-X | None | Economic Impact | Industry | ||
| Victorella pavida (s. l.) is an important fouling organism in power plants and other industrial users of estuarine waters in the Sea of Azov (Partaly 1979). | |||||
| MED-X | None | Ecological Impact | Competition | ||
| 'Thus, larvae of bryozoan foulings settle in the Sea of Azov in the summer and appear in biocoenosis with a high number of mature barnacles and growing hydroids. The bryozoans settle on the remaining free area of barnacles and growing hydroids. The bryozoans settle on the remaining free area of barnacles and hydroids and depress them.' (Partaly 1979). |
|||||
| M130 | Chesapeake Bay | Ecological Impact | Competition | ||
| Victorella pavida (s. l.) is one of the dominant fouling organisms in summer in Chesapeake Bay (Osburn 1944; Cory 1967; Lippson and Lippson 1984; Abbe 1987) and is s presumed to compete with other fouling taxa for space in mesohaline fouling communities, including native barnacles and the introduced hydroids Cordylophora caspia and introduced Garveia franciscana, which are also frequently dominant in fouling communities (Cory 1967; Nauman and Cory 1969; Abbe 1987; Ruiz et al. unpublished data). |
|||||
| M130 | Chesapeake Bay | Ecological Impact | Habitat Change | ||
| Victorella pavida (s. l.) colonies provide habitat for a wide variety of small motile animals (amphipods, polychaetes, etc.) in Chesapeake Bay (Ruiz et al. unpublished observations). | |||||
| M130 | Chesapeake Bay | Economic Impact | Industry | ||
| Victorella pavida (s. l.) is an important fouling organism in power plants and other industrial users of estuarine waters in the Patuxent River, Chesapeake Bay (Nauman and Cory 1969). | |||||
| M130 | Chesapeake Bay | Economic Impact | Shipping/Boating | ||
| Boating - Victorella pavida (s. l.) is 'a nuisance fouling the bottom of boats' in Chesapeake Bay (Osburn 1944; Lippson and Lippson 1984). |
|||||
Regional Distribution Map
Non-native
Native
Cryptogenic
Failed
Occurrence Map
References
Abbe, George R. (1987) Epifauna, In: Heck, Kenneth L.(Eds.) Ecological studies in the middle reach of Chesapeake Bay- Calvert Cliffs. , Berlin. Pp. 82-91Andrews, J. D. (1953) <missing title>, Chesapeake Bay Institute, Johns Hopkins University, Baltimore, Maryland. Pp. <missing location>
Banta, William C.; Backus, Byron T. (1991) <missing title>, US Department of the Interior, Washington DC. Pp. <missing location>
Barnes, Robert D. (1983) Invertebrate Zoology, Saunders, Philadelphia. Pp. 883
Bibbins, Arthur (1892) On the distribution of Cordylophora in the Chesapeake estuaries, and the character of its habitat, Transactions of the Maryland Academy of Sciences 1: 213-228
Bousfield, E. C. (1885) The Victorella pavida of Saville Kent, Annals and Magazine of Natural History 5th series(96): 401-407
Calder, Dale R., Maturo, Jr., J.S. (1978) Phylum Bryozoa, In: Zingmark, Richard G.(Eds.) An Annotated Checklist of the Biota of the Coastal Zone of South Carolina. , Columbia. Pp. 226-229
Calder, Dale Ralph (1966) <missing title>, M.S. Thesis, College of William and Mary, Williamsburg, VA. Pp. <missing location>
Carlton, James T. (1979) History, biogeography, and ecology of the introduced marine and estuarine invertebrates of the Pacific Coast of North America., Ph.D. dissertation, University of California, Davis. Pp. 1-904
Carrada, C. C., Sacchi, C. F. (1964) Recherches ecologiques sur le bryozoaire ctenostome Victorella pavida (Kent)., Vie et Milieu 15: 389-427
Carter, Michelle C.; Bishop, John D. D.; Evans, Nick J.; Wood, Chris A. (2010) Environmental influences on the formation and germination of hibernacula in the brackish-water bryozoan Victorella pavida Saville Kent, 1870 (Ctenostomata: Victorellidae), Journal of Experimental Marine Biology and Ecology 383: 89-95
Cohen, Andrew N. and 10 authors (2005) <missing title>, San Francisco Estuary Institute, Oakland CA. Pp. <missing location>
Cohen, Andrew N.; Carlton, James T. (1995) Nonindigenous aquatic species in a United States estuary: a case study of the biological invasions of the San Francisco Bay and Delta, U.S. Fish and Wildlife Service and National Sea Grant College Program (Connecticut Sea Grant), Washington DC, Silver Spring MD.. Pp. <missing location>
Cory, R. L.; Nauman, J. W. (1969) Epifauna and thermal additions in the upper Patuxent River estuary., Chesapeake Science 10(3-4): 210-217
Cory, Robert L. (1967) Epifauna of the Patuxent River Estuary, Chesapeake Science 8(2): 71-89
Cowles, R.P. (1930) A biological study of the offshore waters of Chesapeake Bay, United States Bureau of Fisheries Bulletin 46: 277-381
D' Hondt, Jean-Loup (1983) Tabular keys for the identification of the recent ctenostomatous Bryozoa, Memoires de l'Institut Oceanographique (Monaco) 14: 1-124
de Rivera, Catherine, and 27 authors (2005) Broad-scale non-indigenous species monitoring along the West Coast in National Marine Sanctuaries and National Estuarine Research Reserves report to National Fish and Wildlife Foundation, National Fish and Wildlife Foundation, Washington, D.C.. Pp. <missing location>
Dean, Thomas A.; Bellis, Vincent J. (1975) Seasonal and spatial distribution of epifauna in the Pamlico River Estuary, North Carolina, Journal of the Elisha Mitchell Scientific Society 91(1): 1-12
Everitt, Betty (1975) Fresh-water Ectoprocta: distribution and ecology of five species in southeastern Louisiana, Transactions of the American Microscopical Society 94(1): 130-134
Goulletquer, Philippe; Bachelet, Guy; Sauriau, Pierre; Noel, Pierre (2002) Invasive aquatic species of Europe: Distribution, impacts, and management, Kluwer Academic Publishers, Dordrecht. Pp. 276-290
Hayward, P.J. (1985) Ctenostome Bryozoans., Synopses of the British Fauna (New Series) 33: 1-169
Humphries, Edythe M.; Duedall, Iver W.; Jordan, Stephen J. (1985) Coal-waste blocks as a fouling substrate in estuarine water., In: (Eds.) Energy Wastes in the Ocean. , New York. Pp. 613-649
Hyman, Libbie H. (1959) <missing title>, McGraw-Hill, New York. Pp. 350-408
Jebram, Diethardt (1980) Laboratory diets and qualitative nutritional requirements for bryozoans, Zoologischer Anzeiger 205: 333-344
Jebram, Diethardt; Everitt, Betty (1982) New victorellids (Bryozoa, Ctenostomata) from North America: the use of parallel cultures in bryozoan taxonomy., Biological Bulletin 163: 172-187
Jung, Kyoung Jin; Wöss, Emmy R.; Chae, Hyun Sook; Seo, Ji Eun (2017) Gymnolaemate bryozoans in fresh and brackish waters of f South Korea: Occurrence, taxonomical temarks and zoogeographical implications, Animal Systematics, Evolution and Diversity 33(1): 37-44
Kasaei, S. Mojtaba; Nasrolahi, Ali; Abtahi, Behrooz; Taylor, Paul D. (2017) Bryozoa of the southern Caspian Sea, Iranian coast, Check List 13(4): 305–313
https://doi.org/10.15560/13.4.305
Larsen, Peter F. (1985) The benthic fauna associated with the oyster reefs of the James River estuary, Virginia, U. S. A., Internationale Revue der Gesamten Hydrobiologie 70(9): 707-814
Lippson, Alice J.; Haire, Michael S.; Holland, A. Frederick; Jacobs, Fred; Jensen, Jorgen; Moran-Johnson, R. Lynn; Polgar, Tibor T.; Richkus, William (1979) Environmental atlas of the Potomac Estuary, Martin Marietta Corp., Baltimore, MD. Pp. <missing location>
Lippson, Alice Jane; Lippson, Robert L. (1984) <missing title>, Johns Hopkins University Press, Baltimore. Pp. <missing location>
Marcus, E. (1955) Notas Sobre Briozoos Marinhos Brasileiros, Arquivos do Museum Nacional Rio de Janeiro 42(1): 273-324
MarLin- Marine Life Information Network 2006-2024 MarLin- Marine Life Information Network. <missing URL>
Massard, Joseph A.; Geimer, Gaby; Bromley, Heather; Dimentman, Chanan (1992) Additional note on the fresh and brackish water Bryozoa of Israel (Phylactolaemata, Gymnolaemata), Bulletin de la Societe des Naturalistes Luxembourgeois 93: 199-214
Maturo, Frank J. S., Jr. (1957) A study of the Bryozoa of Beaufort, North Carolina and vicinity, Journal of the Elisha Mitchell Scientific Society 72(1): 11-68
Menon, N. Ravindranatha; Nair, N. Balakrishnan (1967) Observations on the structure and ecology of Victorella pavida Kent (Bryozoa) from the south west coast of India, Internationale Revue der Gesamten Hydrobiologie 52(3): 237-256
Millar, R. H. (1962) Some Ascidians from the Caribbean, Studies of the Fauna of Curacao and other Caribbean Islands 59: 61-77
Mordukhai-Boltovskoi, Ph. D. (1964) Caspian fauna beyond the Caspian Sea, Internationale Revue der Gesamten Hydrobiologie 49(1): 139-170
Mordukhay-Boltovskoi, Ph. D. (1964) Caspian fauna in fresh waters outside the Ponto Caspian basin, Hydrobiologia 13(1-2): 159-164
Nauman, J. W.; Cory, R. L. (1969) Thermal additions and epifaunal organisms at Chalk Point, Maryland., Chesapeake Science 10(3-4): 218-226
Nehring, Stefan (2001) After the TBT era: alternative anti-fouling paints and their ecological risks., Senckenbergiana Maritima 31(2): 341-351
Occhipinti Ambrogi, Anna (1983) Revue critique des bryozoaires des lagunes italiennes, Commission Internationales pour L'Exploration Scientifique de la Mer Méditerranée, Monaco, Rapports et Procès-Verbaux des Réunions 28(6): 243-244
Occhipinti Ambrogi, Anna; D'Hondt, Jean-Loup (1981) Distribution of bryozoans in brackish waters of Italy., In: Larwood, Gilbert P. and Nielsen, Claus(Eds.) Recent and Fossil Bryozoa.. , Fredensborg. Pp. <missing location>
Olenin, S., Leppakoski, E. 2000-2016 Inventory of Baltic Sea alien species. <missing URL>
Osburn, Raymond C. (1932) Bryozoa from Chesapeake Bay, Ohio Journal of Science 32: 441-447
Osburn, Raymond C. (1944) A survey of the Bryozoa of Chesapeake Bay, Chesapeake Biological Laboratory Publications 63: 1-55
Partaly, E. M. (1979) Ecology of bryozoan foulings in the Sea of Azov, Soviet Journal of Marine Biology 5(3): 213-216
Patrick, Ruth (1994) Rivers of the United States, In: (Eds.) . , New York. Pp. <missing location>
Pfitzenmeyer, Hayes T. (1976) Some effects of salinity on the macroinvertebrates of the lower Potomac., In: Mason, William T.; Flynn, Kevin C.(Eds.) The Potomac Estuary: Biological resources - Trends and Options. , Bethesda MD. Pp. 75-80
Poirrier, Michael A.; Mulino, Maureen M. (1977) Effects of environmental factors on the distribution and morphology of Victorella pavida (Ectoprocta) in Lake Ponchartrain, Louisiana, and vicinity, Chesapeake Science 18(4): 347-352
Poirrier, Michael A.; Mulino, Maureen M. (1977) Impact of the 1975 Bonnet Carré spillway opening on epifaunal invertebrates in southern Lake Pontchartrain, Journal of the Elisha Mitchell Scientific Society 93(1): 11-18
Rao, S. K.; Ganapati, P. N. (1975) Littoral Bryozoa in the Godavary Estuary, Bulletin of the Department of Marine Science of the University of Cochin 7(3): 591-600
Ruiz, Gregory M.; Geller, Jonathan (2018) Spatial and temporal analysis of marine invasions in California, Part II: Humboldt Bay, Marina del Re, Port Hueneme, and San Francisco Bay, Smithsonian Environmental Research Center & Moss Landing Laboratories, Edgewater MD, Moss Landing CA. Pp. <missing location>
Ryland, J. S. (1965) <missing title>, Organization for Economic Co-operation and Development, Paris. Pp. <missing location>
Saville-Kent, William S. (1870) On a new polyzoan, " Victorella pavida," from the Victoria docks., Quarterly Journal of Microscopical Science 10: 35-39
Smithsonian Marine Station at Fort Pierce 2011 Field Guide to the Indian River Lagoon. <missing URL>
Soule, Dorothy F.; Soule, John D.; Morris, Penny A.; Chaney, Henry A (2007) The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon (4th edition), University of California Press, Berkeley CA. Pp. 866-904
Thompson, Michelle Lynne (1993) <missing title>, M.S. Thesis, College of William and Mary, Williamsburg VA. Pp. <missing location>
Turoboyski, K. (1973) Biology and ecology of the crab Rhithropanopeus harrisii ssp. tridentatus, Marine Biology 23: 303--313
Winston, Judith E. (1977) Distribution and ecology of estuarine ectoprocts: a critical review., Chesapeake Science 18(1): 34-57
Winston, Judith E. (1982) Marine bryozoans (Ectoprocta) of the Indian River area (Florida)., Bulletin of the American Museum of Natural History 173: 99-176
Winston, Judith E. (1995) Ectoproct diversity of the Indian River coastal lagoon, Bulletin of Marine Science 57(1): 84-93
Winston, Judith E.; Hayward, Peter J. (2012) The marine bryozoans of the northeast coast of the United States: Maine to Virginia, Virginia Museum of Natural History Memoir 11: 1-180
Wood, Timothy S.; Anurakpongsatorn, Patana Mahujchariyawong, Jukkrit (2006) Freshwater bryozoans of Thailand (Ectoprocta and Entoprocta), Natural History Journal of Chulalongkorn University 6(2): 83-119
Zimmer, Russel L., Reed, Christopher, G. (1994) <missing title>, Johns Hopkins University Press, Baltimore, MD. Pp. 226-246