Invasion History
First Non-native North American Tidal Record: 1993First Non-native West Coast Tidal Record: 1993
First Non-native East/Gulf Coast Tidal Record:
General Invasion History:
Acartiella sinensis is a planktonic calanoid copepod native to estuaries in Southeastern China from Fujian Province to Thailand (Shen and Lee 1963; Chen and Zhang 1965; Orsi and Ohtsuka 1999; Wong et al. 2000). It has also been reported from harbors in Sri Lanka (Chandrasekera and Fernanado 2009) and a freshwater lake in Thailand (Pholpunthin 1997). It is associated with warm, low-salinity waters (Chen and Zhang 1965). In 1993, it was discovered in Suisun Slough, in the San Francisco estuary, where it was introduced through ballast water, and is now abundant (Orsi and Ohtsuka 1999; Bollens et al. 2011). Acartiella sinensis was also collected from the Columbia River estuary in very small numbers, but did not become established (Sytsma et al. 2004).
North American Invasion History:
Invasion History on the West Coast:
Acartiella sinensis was first collected on October 7, 1993 in Suisun Slough and Suisun Bay, in the inland portion of the San Francisco Bay estuary, California. It was subsequently found in much of the estuary, from freshwater to salinities above 20 PSU, but is most abundant in areas from 2.5 to 8.6 PSU (Orsi and Ohtsuka 1999). It occurs in all three sections of the lower estuary (San Pablo, Central, and South San Francisco Bay), but is most abundant in San Pablo Bay in January through May, where and when salinities are likely to be lowest, especially during years of high river flows (Bollens et al. 2011).
Acartiella sinensis has been collected in small numbers in the Columbia River estuary, Oregon starting in 1997, but 'only a few specimens have been found and populations are not known to be established’ (Sytsma et al. 2004). This copepod has been found in ballast water aboard ships in Puget Sound, Washington so future invasions are possible (Cordell et al. 2008a).
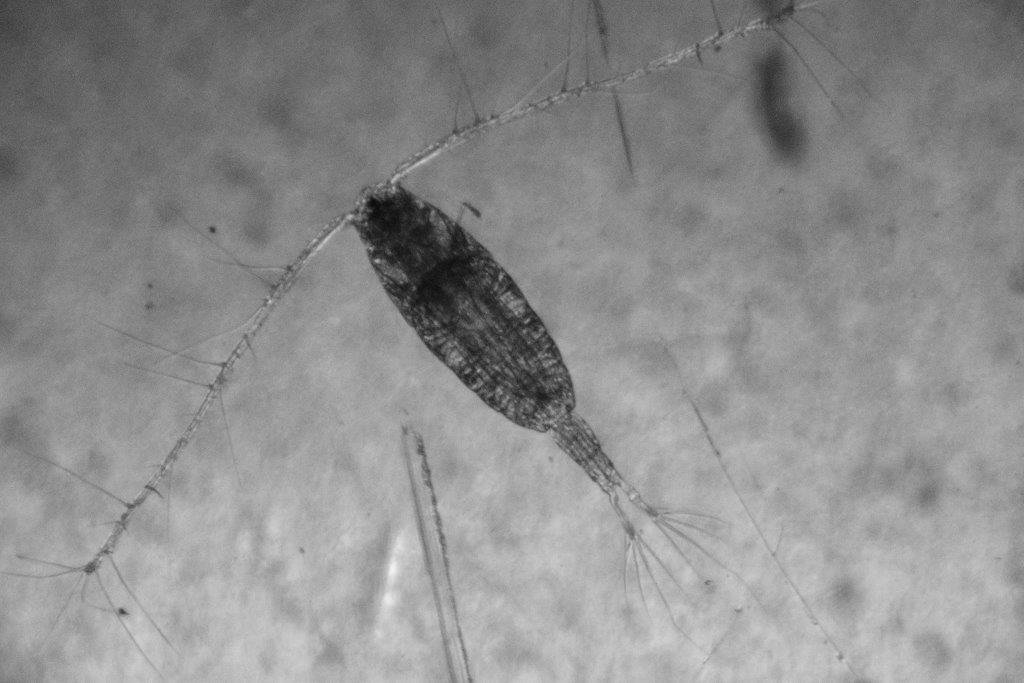
Description
Copepods of the family Acartiidae are characterized by small size, and a slender, elongate body form. The head is separate from the thorax, and has a blunt apex, slightly narrowed distally, with a single eye, lacking cuticular lenses. The posterior corners of the thoracic segments are rounded (Brodskii 1967). The caudal rami are narrow and elongated. This copepod is characteristic of warm, low-salinity water, and sometimes occurs in fresh water (Shen and Lee 1963).
In female Acartiella sinensis, the 5th legs are biramous, and 2-segmented, with the exopod slender and spine-like. The anal somite of the urosome is fused with the caudal rami. The caudal rami in the female are asymmetrical, bending somewhat to the left (in dorsal view). Urosome segments 1 and 2 are fused in the female, with the genital openings occurring in the center of the ventral area of the segment (Shen and Lee 1963; Chen and Zhang 1965; Barthelemy 1999). Adult females in San Francisco Bay were 1.27-1.64 mm long (Orsi and Ohtsuka 1999), comparable to a length of 1.38 mm in China (Shen and Lee 1963).
In male A. sinensis, the right (in dorsal view) antennule (1st antenna) of the male is prehensile, with segments 11-15 thickened and the antennule hinged between segments 16 and 17. The 5th legs in the male are asymmetrical, and comprised of 3 segments each (Shen and Lee 1963; Chen and Zhang 1965). Males were 1.17-1.34 mm long in San Francisco Bay (Orsi and Ohtsuka 1999) and 1.23 mm long in China (Shen and Lee 1963).
To our knowledge, naupliar and copepodite development in this genus has not been described. Nauplii probably resemble those of Acartia spp.
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Arthropoda | |
| Subphylum: | Crustacea | |
| Class: | Maxillopoda | |
| Subclass: | Copepoda | |
| Order: | Calanoida | |
| Family: | Acartiidae | |
| Genus: | Acartiella | |
| Species: | sinensis |
Synonyms
Potentially Misidentified Species
Copepods of 'Acartia clausi species group, species name uncertain (Bollens et al. 2011)
Acartia californiensis
Native, estuarine form (Bollens et al. 2011)
Acartiella gravelyi
Indian Ocean (Razouls et al. 2012)
Acartiella kempi
Indian Ocean
Acartiella keralensis
Indian Ocean
Acartiella major
Indian Ocean
Acartiella minor
Indian Ocean
Acartiella nicolae
Indonesia
Acartiella sewelli
Indian Ocean
Acartiella tortaniformis
Indian Ocean
Ecology
General:
Planktonic calanoid copepods mate in the water column. Males use their modified antenules and 5th pair of swimming legs to grasp the female and transfer spermatophores to the female's genital segment. Female copepods of the family Acartiidae lay unattached eggs, one at a time, in the water column (Barnes 1983). Eggs hatch into nauplii which go through six stages. The first stage, NI, has 3 pairs of appendages and is unsegmented - each molt has additional appendages and/or more differentiation of segments. The sixth stage (NVI) molts into a first copepodite stage (CI), with the basic form of the adult, and fully differentiated feeding structures, but with only two pairs of swimming legs, and only one urosomal segment. The copepod goes through five additional molts, with increasing numbers of swimming legs, urosomal segments, and sexual differentiation. The sixth (CVI) stage is the male or female adult (Barnes 1983; Johnson and Allen 2005).
Acartiella sinensis is characteristic of estuaries with warm, low-salinity waters (Chen and Lee 1963; Orsi and Ohtsuka 1999; Wong et al. 2000), and has landlocked freshwater resident populations (Pholpuntin 1997). Early stages probably feed on phytoplankton, but adults also capture copepod nauplii and copepodites (Barnes 1983). In experiments, adults of A. sinensis showed little feeding on protists, but did feed on early stages of Limnoithona tetraspina (York et al. 2014). In later studies, Acartiella sinensis was found to have a significant predation impact on nauplii of Pseudodaptomus forbesi and Limnoithona tetraspina, averaging 11 and 4 % per day, respectively. Rates of predation on adults and copepodites were ~1% per day (Slaughter et al. 2016).
Food:
Copepod nauplii; phytoplankton
Consumers:
Mysids, fishes
Trophic Status:
Omnivore
OmniHabitats
| General Habitat | Unstructured Bottom | None |
| Salinity Range | Limnetic | 0-0.5 PSU |
| Salinity Range | Oligohaline | 0.5-5 PSU |
| Salinity Range | Mesohaline | 5-18 PSU |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Planktonic | None |
Tolerances and Life History Parameters
| Minimum Temperature (ºC) | 9 | Field, San Francisco Bay (Bollens et al. 2011) |
| Maximum Temperature (ºC) | 29 | Field, Zhujiang River (Pearl River) estuary, China (Wong et al. 2000) |
| Minimum Salinity (‰) | 0 | Lower limit for abundant occurrence in San Francisco estuary 2.5 PSU (Orsi and Ohtsuka 1999). |
| Maximum Salinity (‰) | 20 | Upper limit for abundant occurrence in San Francisco estuary, 8.6 ppt (Orsi and Ohtsuka 1999). |
| Minimum Length (mm) | 1.2 | Minimum adult length, males, for females 1.27 (San Francisco Bay (Orsi and Ohtsuka 1999). |
| Maximum Length (mm) | 1.6 | Maximum |
| Broad Temperature Range | None | Warm temperate-Tropical |
| Broad Salinity Range | None | Oligohaline-Polyhaline |
General Impacts
The planktonic copepod Acartiella sinensis is abundant, but not dominant in the upstream portions of the San Francisco estuary (Orsi and Ohtsuka 1999; Bollens et al. 2011). The extent to which is has displaced native species, or altered food webs, in the estuary is unclear. In experiments, A. sinensis fed heavily on nauplii and copepodites of Limnoithona tetraspina and Pseudodiaptomus forbesi (York et al. 2014; Slaughter et al. 2016).Regional Impacts
| NEP-V | Northern California to Mid Channel Islands | Ecological Impact | Predation | ||
| Acartiella sinensis had a significant predation impact on nauplii of Pseudodaptomus forbesi and Limnoithona tetraspina, averaging 11 and 4 % per day, respectively. Rates of predation on adults and copepodites were ~1% per day (Slaughter et al. 2016). | |||||
| P090 | San Francisco Bay | Ecological Impact | Predation | ||
| Acartiella sinensis had a significant predation impact on nauplii of Pseudodaptomus forbesi and Limnoithona tetraspina, averaging 11 and 4 % per day, respectively. Rates of predation on adults and copepodites were ~1% per day (Slaughter et al. 2016). | |||||
| CA | California | Ecological Impact | Predation | ||
| Acartiella sinensis had a significant predation impact on nauplii of Pseudodaptomus forbesi and Limnoithona tetraspina, averaging 11 and 4 % per day, respectively. Rates of predation on adults and copepodites were ~1% per day (Slaughter et al. 2016)., Acartiella sinensis had a significant predation impact on nauplii of Pseudodaptomus forbesi and Limnoithona tetraspina, averaging 11 and 4 % per day, respectively. Rates of predation on adults and copepodites were ~1% per day (Slaughter et al. 2016). | |||||
Regional Distribution Map
Non-native
Native
Cryptogenic
Failed
Occurrence Map
References
Barnes, Robert D. (1983) Invertebrate Zoology, Saunders, Philadelphia. Pp. 883Barthelemy, R-M. (1999) Functional morphology and taxonomic relevance of the female genital structures in Acartiidae (Copepoda: Calanoida), Journal of the Marine Biological Association of the United Kingdom 79: 857-870
Bollens, Stephen M.; Breckenridge, Joanne K.; Cordell, Jeffery R. Simenstad, Charles A.; Kalata, Olga (2014) Zooplankton of tidal marsh channels in relation to environmental variables in the upper San Francisco Estuary, Aquatic Biology 21: 205-219
Bollens, Stephen M.; Breckenridge, Joanne K. Vanden Hoof, Rian C.; Cordell, Jeffery R. (2011) Mesozooplankton of the lower San Francisco Estuary: spatio-temporal patterns, ENSO effects and the prevalence of non-indigenous species, Journal of Plankton Research 33(9): 1358-1377
Brodskii, K. A. (1967) <missing title>, !srael Program for Scientific Translations, Jerusalem, Israel. Pp. <missing location>
Chandrasekera, W. U.; Fernando, M. A. S. T. (2009) Accidental introduction of alien plankton into the Sri Lankan coastal zone through ballast water of cargo ships, Sri Lanka Journal of Aquatic Science 14: 87-103
Chen, Q.; Zhang, S. (1965) The planktonic copepods of the Yellow Sea and East China Sea I. Calanoida, Studia Marina Sinica 7: 20-31
Cohen, Andrew N.; Carlton, James T. (1995) Nonindigenous aquatic species in a United States estuary: a case study of the biological invasions of the San Francisco Bay and Delta, U.S. Fish and Wildlife Service and National Sea Grant College Program (Connecticut Sea Grant), Washington DC, Silver Spring MD.. Pp. <missing location>
Cordell, Jeffrey R. and 5 authors (2009) Factors influencing densities of non-indigenous species in the ballast water of ships arriving at ports in Puget Sound, Washington, United States, Aquatic Conservation: Marine and Freshwater Ecosystems 19(3): 322-343
Cordell, Jeffrey; Bollens, Stephen M.; Draheim, Robyn; Sytsma, Mark (2008) Asian copepods on the move: Recent invasions in the Columbia-Snake River system, USA, ICES Journal of Marine Science 65: 753-758
Orsi, James J., Ohtsuka, Susumu (1999) Introduction of the Asian copepods Acartiella sinensis, Tortanus dextrilobatus (Copepoda:Calanoida), and Limnoithona tetraspina (Copepoda: Cyclopoida) to the San Francisco Estuary,California, USA., Plankton Biology and Ecology 46(2): 128-131
Pholpuntin, P. (1997) Freshwater zooplankton (Rotifera, Cladocera, and Copepoda) from Thale-noi, south Thailand, Journal of the Science Society of Thailand 23(1): 23-33
Razouls, C.; de Bovée, F.; Kouwenberg, J.Desreumaux, N. 2005-2012 Diversity and geographic distribution of marine planktonic copepods. <missing URL>
Ruiz, Gregory; Geller, Jonathan (2021) Spatial and temporal analysis of marine invasions: supplemental studies to evaluate detection through quantitative and molecular methodologies, Marine Invasive Species Program, California Department of Fish and Wildlife, Sacramento CA. Pp. 153 ppl.
Shen, C-J.; Lee, F.-S. (1963) [The estuarine copepods of Chiekong and Zaikong Rivers, Kwangtung Province, China] (Chinese), Studia Marina Sinica 3: 571-596
Slaughter, Anne M.; Ignoffo, Toni R.; Kimmerer, Wim (2016) Predation impact of Acartiella sinensis, an introduced predatory copepod in the San Francisco Estuary, USA, Marine Ecology Progress Series 547: 47-60
Sytsma, Mark D.; Cordell, Jeffrey R.; Chapman, John W.; Draheim, Robyn, C. (2004) <missing title>, Center for Lakes and Reservoirs, Portland State University, Portland OR. Pp. <missing location>
Winder, Monika; Jassby, Alan D. (2011) Shifts in zooplankton community structure: implications for food web processes in the upper San Francisco estuary, Estuaries and Coasts 34: 675-690
Winder; Monika; Jassby, Alan D.; Mac Nally, Ralph (2011) Synergies between climate anomalies and hydrological modifications facilitate estuarine biotic invasions, Ecology Letters 14: 749-757
Wong, C, K,; Tam, P. F.; Fu, Y. Y.; Chen, Q. C. (2000) The biodiversity crisis and Crustacea: proceedings of the Fourth International Crustacean Congress, Balkema, Amsterdam. Pp. 363-379
York, Joanna K.; McManus, George B.; Kimmerer, Wim J.; Slaughter, Anne M.; Ignoffo, Toni R. (2014) Trophic links in the plankton in the low salinity zone of a large temperate estuary: Top-down effects of introduced copepods, Estuaries and Coasts 37: 576-588