Invasion History
First Non-native North American Tidal Record: 1993First Non-native West Coast Tidal Record: 1993
First Non-native East/Gulf Coast Tidal Record:
General Invasion History:
Hyperacanthomysis longirostris is native to coastal waters of southern Japan, Korea, and China (Modlin and Orsi 1997; Huang 2001; Fukuoka and Murano 2005). In at least one estuarine system, the Chikugo River estuary, Ariake Sea, Japan, this mysid prefers low salinities (0-10 PSU, Suzuki et al. 2009). It has been introduced to the Sacramento-San Joaquin Delta, California.
North American Invasion History:
Invasion History on the West Coast:
In July 1993, an unidentified mysid was caught off Sherman Island, near Antioch, California in the Sacramento-San Joaquin Delta. It was identified as a new species, Acanthomysis bowmani (Modlin and Orsi 1997). NB: This name is still used extensively in the San Francisco estuary literature. Modlin and Orsi noted its similarity to the mysid then known as A. longirostris. Fukuoka and Murano (2005) synonymized bowmani with longirostris, and placed longirostris in a new genus, Hyperacanthomysis. Hyperacanthomysis longirostris is abundant throughout the Delta and Suisun Bay, and reaches its peak abundance at about 2 PSU (Modlin and Orsi 1997). By 1995, H. longirostris became the predominant mysid in the Delta, largely replacing the native Neomysis mercedis (Baxter and Heib 2006; Nobriga and Feyrer 2008). It appears to be rare in the lower estuary, but was collected once at China Camp marsh, San Pablo Bay, in May 2002 (Dean et al. 2005).
Description
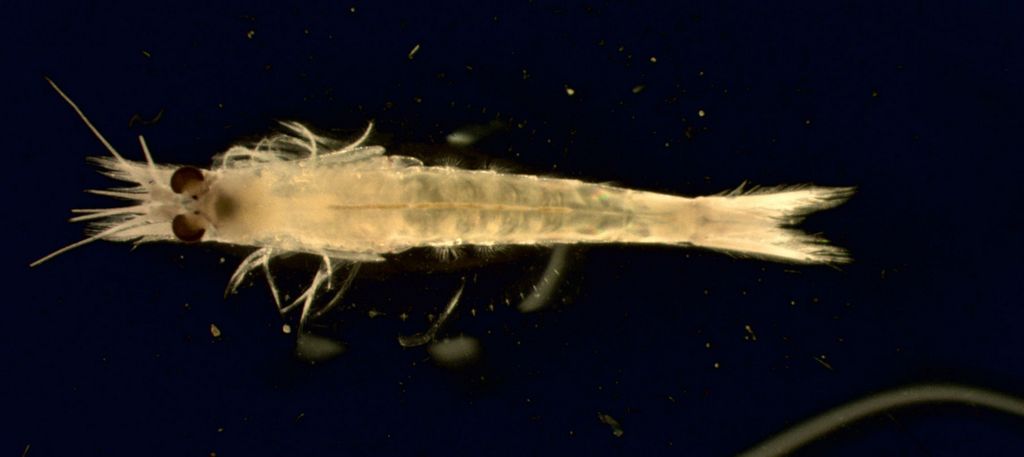
The estuarine mysid Hyperacanthomysis longirostris was originally described as Acanthomysis longirostris from Japan in 1936 (Fukuoka and Murano 2005). It was described as a new species, A. bowmani, after it was discovered in the San Francisco estuary in 1993 (Modlin and Orsi 1997). This name is still used extensively in the San Francisco estuary literature. Modlin and Orsi (1997) noted its similarity to the mysid then known as A. longirostris. Fukuoka and Murano (2005) synonymized A. bowmani with longirostris, and placed longirostris in a new genus, Hyperacanthomysis.
Mysids are small, shrimplike crustaceans, with a cape-like carapace which covers the head and thorax, but which is not attached to the last four thoracic segments. A pair of spherical statocysts is located at the base of the inner uropods. Adult females have a ventral brood pouch. Hyperacanthomysis longirostris has a slender, elongated body. The anterior edge of the carapace is produced into a sharp, triangular rostrum, with keel and struts at the base. The posterior margin of the carapace has a curved indentation, exposing somites 7 and 8. The eyes are stalked and prominent with oval corneas. The abdominal segments are smooth, without furrows or folds. The distal tip of the antennal scale is rounded. In the male, the basipod of the 7th thoracic leg is modified into a penis, about 2.4 X as long as wide. The exopods and endopods of the uropods are blade-like, with the lateral margin slightly concave. The endopods each have 2 spines between statocysts and the margin. The telson is tongue-like, 2.4 X as long as wide, with the lateral margin of telson completely armed with 18-19 spines, and 4 terminal spines that are 3.3 X longer than marginal spines. Adult males are 7.7-11.8 mm and females are 10.5-11.8 mm in size. Description based on Modlin and Orsi 1997, Fukuoka and Murano 2000, and Modlin, in Carlton 2007.
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Arthropoda | |
| Subphylum: | Crustacea | |
| Class: | Malacostraca | |
| Subclass: | Eumalacostraca | |
| Superorder: | Peracarida | |
| Order: | Mysidacea | |
| Suborder: | Mysida | |
| Family: | Mysidae | |
| Genus: | Hyperacanthomysis | |
| Species: | longirostris |
Synonyms
Acanthomysis longirostris (Ii, 1936)
Potentially Misidentified Species
Single spine in vicinity of statocyst, found with H. longirostris in low-salinity zone
Orientomysis hwanhaiensis
Four spines in vicinity of statocyst, occurs in lower estuary in polyhaline zone
Ecology
General:
Mysids are small, shrimplike crustaceans, which tend to divide their time between epibenthic habitats and the plankton, sometimes migrating diurnally, or else making brief swims between benthic habitats. Males have a pair of penes located at the junction of the thorax and abdomen, and modified pleopods used for the transfer of spermatophores, which in H. longirostris is pleopod 4. Adult females have a brood pouch, and give birth to juveniles, resembling miniature adults (Barnes 1983; Bowman and Orsi 1993; Modlin, in Carlton 2007).
Hyperacanthomysis longirostris is concentrated in low-salinity estuarine habitats in the upper San Francisco estuary. It maintains its population in 0-5 PSU, largely by a pattern of vertical migration, migrating upward on incoming tides (Kimmerer et al. 1998). In a Japanese estuary, it also showed a pattern of preference for low salinities, with most of the population occurring between 0 and 10 PSU (Suzuki et al. 2009). Adult females migrated upstream, presumably using tidal currents, above the 1 PSU front, where females gave birth to juveniles, who drifted downstream as they grew (Suzuki et al. 2009). Mysids may be epibenthic feeders, suspension feeders, predators, or omnivores (Barnes 1983). Specific information on feeding of H. longirostris is not available, but given its abundance, omnivorous feeding is likely.
Food:
Zooplankton; Phytoplankton
Consumers:
Fishes, decapod shrimps
Trophic Status:
Omnivore
OmniHabitats
| General Habitat | Tidal Fresh Marsh | None |
| General Habitat | Salt-brackish marsh | None |
| General Habitat | Unstructured Bottom | None |
| Salinity Range | Limnetic | 0-0.5 PSU |
| Salinity Range | Oligohaline | 0.5-5 PSU |
| Salinity Range | Mesohaline | 5-18 PSU |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Planktonic | None |
| Vertical Habitat | Nektonic | None |
Life History
Tolerances and Life History Parameters
| Minimum Temperature (ºC) | 8 | Suzuki et al. 2009, Chikugo River estuary, (Suzuki et al. 2009) |
| Maximum Temperature (ºC) | 30 | Suzuki et al. 2009, Chikugo River estuary, (Suzuki et al. 2009) |
| Minimum Salinity (‰) | 0.5 | Ranges into tidal freshwater part of Delta (Modlin and Orsi 1997) |
| Maximum Salinity (‰) | 22 | Field, China Camp marsh, 5/2002, Dean et al. 2005; most abundant at 2 PSU (Modlin and Orsi 1997) |
| Minimum Length (mm) | 7.7 | Adults (Modlin and Orsi 1997) |
| Maximum Length (mm) | 11.8 | Adults (Modlin and Orsi 1997) |
| Broad Temperature Range | None | Cold temperate-Warm temeprate |
| Broad Salinity Range | None | Tidal Limnetic-Mesohaline |
General Impacts
The mysid Hyperacanthomysis longirostrishas largely replaced the native Neomysis mercedis, an important food organism for juvenile fishes, including the Striped Bass Morone saxatilis which is an important sport fish in the San Francisco estuary. This mysid is less abundant than the native, and is smaller, so it has potential economical implications for fisheries in the estuary. However, changes in the mysid community are only one of many changes in the environment and food webs of the estuary, some of which are traceable to the invasion of the Asian clam Corbula amurensis, which led to a shift from a plankton - to a benthos- dominated ecosystem (Winder and Jassby 2011).
Competition: By 1995, H. longirostris became the predominant mysid in the Delta, largely replacing the native Neomysis mercedis (Nobriga and Feyrer 2008). The causes for this replacement have not been examined, but are probably associated with the alteration of San Francisco estuary's food web by the filter-feeding Asian brackish water clam Corbula amurensis.
Food/Prey: Hyperacanthomysis longirostris is smaller than the native Neomysis mercedis, which had been a major food item for juvenile Striped Bass (Morone saxatilis), a major introduced sport fish in the San Francisco estuary. It also maintains smaller population densities, probably due to the reorganization of the estuaries food web by massive populations of the filter-feeding Asian brackish-water clam Corbula amurensis (Feyrer et al. 2003; Nobriga and Feyrer 2008). Feyrer et al. (2003) found that following the clam and exotic mysid invasion, the percentage of mysids in the diet dropped drastically for Striped Bass (spring 90- fall 80% to 40%, post-clam, exotic mysids), Starry Flounder (Platichthys flesus, native) (70-40% to near 0), and Yellowfin Goby (Acanthomysis flavimanus, introduced) (50-20% to 5%). For Striped Bass, the decline of the native mysid N. mercedis and its replacement by the smaller H. longirostris resulted in the fish shifting to piscivory at a smaller size (Feyrer et al. 2003). The decline of the native mysid N. mercedis, and its replacement by smaller numbers of a smaller species may be a major factor in the decline of Striped Bass populations in the estuary (Nobriga and Feyrer 2008; Winder and Jassby 2011).
Regional Impacts
| P090 | San Francisco Bay | Ecological Impact | Competition | ||
| By 1995, H. longirostris became the predominant mysid in the Delta, largely replacing the native Neomysis mercedis (Nobriga and Feyrer 2008). | |||||
| P090 | San Francisco Bay | Ecological Impact | Food/Prey | ||
| Hyperacanthomysis longirostris is smaller than the native Neomysis mercedis, which had been a major food item for juvenile Striped Bass Morone saxatilis, a major introduced sport fish in the San Francisco estuary. It also maintains smaller population densities, probably due to the reorganization of the estuaries foodweb by massive populations of the filter-feeding Asian Brackish-Water Clam Corbula amurensis (Feyrer et al. 2003; Nobriga and Feyrer 2008). Feyrer et al. (2003) found that following the clam and exotic mysid invasion, the percentage of mysids in the diet dropped drastically for Striped Bass (spring 90- fall 80% to 40%, post-clam, exotic mysids), Starry Flounder (Platichthys flesus, native) (70-40% to near 0), and Yellowfin Goby (Acanthomysis flavimanus, introduced) (50-20% to 5%). For Striped Bass, the decline of the native mysid N. mercedis and its relacement by the smaller H. longirostris resulted in the fish starting mysid feeding at a smaller size and shifting to piscivory at a smaller size (Feyrer et al. 2003). The decline of the native mysid, and its replacement by smaller numbers of a smaller species may be a major factor in the decline of Striped Bass populations in the estuary (Nobriga and Feyrer 2008) | |||||
| NEP-V | Northern California to Mid Channel Islands | Ecological Impact | Competition | ||
| By 1995, H. longirostris became the predominant mysid in the Delta, largely replacing the native Neomysis mercedis (Nobriga and Feyrer 2008). | |||||
| NEP-V | Northern California to Mid Channel Islands | Ecological Impact | Food/Prey | ||
| Hyperacanthomysis longirostris is smaller than the native Neomysis mercedis, which had been a major food item for juvenile Striped Bass Morone saxatilis, a major introduced sport fish in the San Francisco estuary (Bryant and Arnold 2007). It also maintains smaller population densities (Baxter et al. 2005), probably due to the reorganization of the estuaries foodweb by massive populations of the filter-feeding Asian Brackish-Water Clam Corbula amurensis (Feyrer et al. 2003; Nobriga and Feyrer 2008). Feyrer et al. (2003) found that following the clam and exotic mysid invasion, the percentage of mysids in the diet dropped drastically for Striped Bass (spring 90- fall 80% to 40%, post-clam, exotic mysids), Starry Flounder (Platichthys flesus, native) (70-40% to near 0), and Yellowfin Goby (Acanthomysis flavimanus, introduced) (50-20% to 5%). For Striped Bass, the decline of the native mysid N. mercedis and its relacement by the smaller H. longirostris resulted in the fish starting mysid feeding at a smaller size and shifting to piscivory at a smaller size (Feyrer et al. 2003). The decline of the native mysid, and its replacement by smaller numbers of a smaller species may be a major factor in the decline of Striped Bass populations in the estuary (Nobriga and Feyrer 2008) | |||||
| CA | California | Ecological Impact | Competition | ||
| By 1995, H. longirostris became the predominant mysid in the Delta, largely replacing the native Neomysis mercedis (Nobriga and Feyrer 2008)., By 1995, H. longirostris became the predominant mysid in the Delta, largely replacing the native Neomysis mercedis (Nobriga and Feyrer 2008). | |||||
| CA | California | Ecological Impact | Food/Prey | ||
| Hyperacanthomysis longirostris is smaller than the native Neomysis mercedis, which had been a major food item for juvenile Striped Bass Morone saxatilis, a major introduced sport fish in the San Francisco estuary (Bryant and Arnold 2007). It also maintains smaller population densities (Baxter et al. 2005), probably due to the reorganization of the estuaries foodweb by massive populations of the filter-feeding Asian Brackish-Water Clam Corbula amurensis (Feyrer et al. 2003; Nobriga and Feyrer 2008). Feyrer et al. (2003) found that following the clam and exotic mysid invasion, the percentage of mysids in the diet dropped drastically for Striped Bass (spring 90- fall 80% to 40%, post-clam, exotic mysids), Starry Flounder (Platichthys flesus, native) (70-40% to near 0), and Yellowfin Goby (Acanthomysis flavimanus, introduced) (50-20% to 5%). For Striped Bass, the decline of the native mysid N. mercedis and its relacement by the smaller H. longirostris resulted in the fish starting mysid feeding at a smaller size and shifting to piscivory at a smaller size (Feyrer et al. 2003). The decline of the native mysid, and its replacement by smaller numbers of a smaller species may be a major factor in the decline of Striped Bass populations in the estuary (Nobriga and Feyrer 2008), Hyperacanthomysis longirostris is smaller than the native Neomysis mercedis, which had been a major food item for juvenile Striped Bass Morone saxatilis, a major introduced sport fish in the San Francisco estuary. It also maintains smaller population densities, probably due to the reorganization of the estuaries foodweb by massive populations of the filter-feeding Asian Brackish-Water Clam Corbula amurensis (Feyrer et al. 2003; Nobriga and Feyrer 2008). Feyrer et al. (2003) found that following the clam and exotic mysid invasion, the percentage of mysids in the diet dropped drastically for Striped Bass (spring 90- fall 80% to 40%, post-clam, exotic mysids), Starry Flounder (Platichthys flesus, native) (70-40% to near 0), and Yellowfin Goby (Acanthomysis flavimanus, introduced) (50-20% to 5%). For Striped Bass, the decline of the native mysid N. mercedis and its relacement by the smaller H. longirostris resulted in the fish starting mysid feeding at a smaller size and shifting to piscivory at a smaller size (Feyrer et al. 2003). The decline of the native mysid, and its replacement by smaller numbers of a smaller species may be a major factor in the decline of Striped Bass populations in the estuary (Nobriga and Feyrer 2008) | |||||
Regional Distribution Map
Non-native
Native
Cryptogenic
Failed
Occurrence Map
References
Barnes, Robert D. (1983) Invertebrate Zoology, Saunders, Philadelphia. Pp. 883Baxter, Randall; Heib, Katherine (2006) Zooplankton Monitoring 2005, IEP Newsletter 19(2): 13-16
Carlton, James T. (Ed.) (2007) The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon Fourth Edition, Completely Revised and Expanded, University of California Press, Berkeley. Pp. <missing location>
Dean, Amy F.; Bollens, Stephen M.; Simenstad, Charles; Cordell, Jeffery (2005) Marshes as sources or sinks of an estuarine mysid: demographic patterns and tidal flux of Neomysis kadiakensis at China Camp marsh, San Francisco estuary, Estuarine, Coastal and Shelf Science 63: 1-11
Feyrer, Frederick; Herbold, Bruce; Matern, Scott A.; Moyle, Peter (2003) Dietary shifts in a stressed fish assemblage: consequences of a bivalve invasion in the San Francisco estuary., Environmental Biology of Fishes 67: 277-288
Fukuoka, Kouki; Murano, Masaaki (2000) Hyperacanthomysis, a new genus Acanthomysis longirostris li, 1936, and A brevirostris Wang & Liu, 1997 (Crustacea: Mysidacea: Mysidae), Plankton Biology and Ecology 47(2): 122-128
Fukuoka, Kouki; Murano, Masaaki (2005) A revision of East Asian Acanthomysis (Crustacea: Mysida: Mysidae) and redefinition of Orientomysis, with description of a new species., Journal of Natural History 39(9): 657-708
Huang, Zongguo (Ed.), Junda Lin (Translator) (2001) Marine Species and Their Distributions in China's Seas, Krieger, Malabar, FL. Pp. <missing location>
Kimmerer, William J.; Burau, J. R.; Bennett, W. A. (1998) Tidally oriented vertical migration and position maintenance of zooplankton in a temperate estuary., Limnology and Oceanography 43(7): 1697-1709
Mecum, W. Lee (2006) <missing title>, California Department of Fish and Game, Sacramento CA. Pp. 1-6
Modlin, Richard F. (2007) The Light and Smith Manual: Intertidal invertebrates from Central California to Oregon (4th edition), University of California, Berkeley CA. Pp. 489-485
Modlin, Richard F.; Orsi, James J. (1997) Acanthomysis bowmani, a new species, and A. aspera Li, Mysidacea newly reported from the Sacramento-San Joaquin Estuary, Califronia (Crustacea: Mysidae), Proceedings of the Biological Society of Washington. 110(3): 439-446
Musey, Larry () Salinity Tolerance of the African Pipid Frog, Xenopus laevis, Copeia 5(2): 584-586
Nobriga, Matthew L.; Feyrer, Frederick (2008) Diet composition in San Francisco Estuary striped bass: does trophic adaptability have its limits?, Environmental Biology of Fishes 83(4): 495-503
Robertson, D. Ross; Dominguez-Dominguez, Omar; Solís-Guzmán; María Gloria; Kingon, Kelly C (2021b) Origins of isolated populations of an Indo-Pacific damselfish at opposite ends of the Greater Caribbean, Aquatic Invasions 16: 269-280
0, https://doi.org/10. 3391/ai.2021.16.2.04 Received: 13 May 20
Suzuki, Keita W.; Kasai, Akihide; Isoda,Takane; Nakayama, Kouji; Tanaka, Masaru (2009) Horizontal distribution and population dynamics of the dominant mysid Hyperacanthomysis longirostris along a temperate macrotidal estuary (Chikugo River estuary, Japan), Estuarine, Coastal and Shelf Science 83: 517-528
Winder, Monika; Jassby, Alan D. (2011) Shifts in zooplankton community structure: implications for food web processes in the upper San Francisco estuary, Estuaries and Coasts 34: 675-690
Winder; Monika; Jassby, Alan D.; Mac Nally, Ralph (2011) Synergies between climate anomalies and hydrological modifications facilitate estuarine biotic invasions, Ecology Letters 14: 749-757