Invasion History
First Non-native North American Tidal Record: 2001First Non-native West Coast Tidal Record: 2001
First Non-native East/Gulf Coast Tidal Record:
General Invasion History:
Epigamia noroi is a syllid polychaete, native to the Northwest Pacific. It is known to range from southern Hokkaido to the Seto Inland Sea, and the Sea of Japan (Imajima 1966b). It is found in the intertidal to 40 m depth living among rocks and seaweed. It has been found in southern California, at La Jolla and Catalina Island (Nygren 2004).
North American Invasion History:
Invasion History on the West Coast:
In January 2001, eight specimens of Epigamia noroi were found in an intertidal Surfgrass (Phyllospadix spp.) bed at La Jolla, California. An additional 31 individuals were found on a floating dock at the Wrigley Marine Science Center on Santa Catalina Island, among hydroids, sponges, and bryozoans (Nygren 2004). These populations are considered to be established (Leslie Harris, personal communication). This worm could have been transported in ship fouling or in ballast water. Epitokous females, of this subfamily, Autolytinae, are often seen in plankton samples, carrying sacs of eggs (Pettibone 1963).
Description
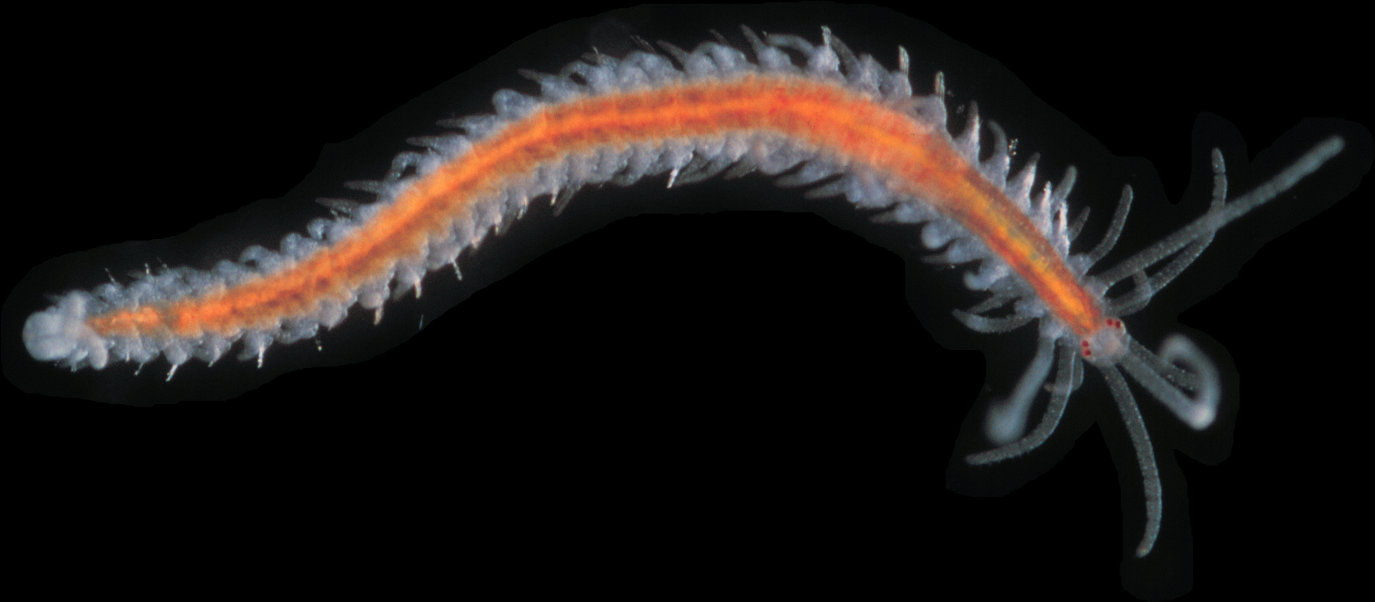
Epigamia noroi is a syllid polychaete. Family characteristics of the Syllidae include: a relatively small and slender body; a prostomium with two pairs of eyes and sometimes a couple of eyespots; three antennae and paired palps; an eversible pharynx and proventricle; the anterior-most segment (peristomium) lacking chaetae, but bearing 1-2 pairs of tentacles; uniramous parapodia (biramous in reproductive individuals); dorsal and ventral cirri on each chaetiger; and a pygidium (anal segment) with 2-3 anal cirri (Pettibone 1963; Blake and Ruff 2007).
During its atokous (non-reproductive) stage, Epigamia noroi is slender and uniform in width, but tapers posteriorly. The prostomium is a compressed ovoid, wider than long, with four eyes arranged in a trapezoid, with the anterior eyes about twice as large as the posterior ones. The prostomium bears three antennae, with the medial one rising in the center of the prostomium. The median antenna extends back to the 8th-12th chaetiger. The lateral antennae are about 2/3 the length of the median antenna. The palps are fused along the median line. The nuchal epaulettes extend to the middle of the 2nd chaetiger. The pharynx is surrounded by 9 large soft papillae, and is S-shaped and convoluted, a feature that, together with the absence of the ventral cirri, characterizes the Autolytinae subfamily. The mouth opening has 9 large and 18 small teeth in a series, so that two small ones alternate with each of the large teeth (Imajima 1966b; Nygren 2004).
The anterior-most segment (peristomium) bears two pairs of dorsal tentacular cirri, also about 2/3 of the length of the median antennae. The ventral tentacular cirri are 1/3-1/2 as long as the dorsal pair. On the first chaetiger, the first dorsal cirri are as long as the median antenna, and the second dorsal cirri as long as the ventral tentacular cirri. The proventricle is ellipsoid and extends from chaetigers 11 to 13 -14. The first dorsal cirri are longer than the median antenna; the second are 1/3 as long, and the subsequent dorsal cirri are about 1/2 to 2/3 of the body width. A band of small cilia extends around each segment. The parapodial lobes are bluntly conical and rounded, with a bundle of chaetae. The pygidium has two long cirri (Imajima 1966b; Nygren 2004). Adults reach 13 mm and 87 chaetigers in length (Imajima 1966b), but Nygren's (2004) specimens were 2.1-6.3 mm and 22-53 chaetigers. The worm is light orange, or sometimes pale bluish with a bright yellow line running down the median portion of the body (Imajima 1966b; Nygren 2004).
Epitokous males have greatly enlarged red eyes. The body is divided into three sections, an anterior section with 11 uniramous chaetigers, a median section with 37 biramous chaetigers, and a posterior region with three uniramous chaetigers. The lateral antennae are bifid (2 branches). The pharynx and proventriculus degenerate. A live male epitoke was 6.3 mm long, with 51 chaetigers. The color pattern was similar to that seen in atokous animals, except that the median yellow line was less distinct in region b (Nygren 2004).
Epitokous females have red eyes. Based on a partially transformed female and on other species in the genus, the body is divided into three sections, an anterior section with ~14 uniramous chaetigers, a median section with 20-41 biramous chaetigers, and a posterior region with 16-33 uniramous chaetigers. The pharynx and proventricle degenerate (Nygren 2004). Eggs in this subfamily (Autloytinae) are carried within the body, or in a large external sac, around which the body is curled (Pettibone 1963; Nygren 2004).
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Annelida | |
| Class: | Polychaeta | |
| Subclass: | Palpata | |
| Order: | Aciculata | |
| Suborder: | Phyllodocida | |
| Family: | Syllidae | |
| SubFamily: | Autolytinae | |
| Genus: | Epigamia | |
| Species: | noroi |
Synonyms
Autolytus nipponensis (Imajima & Hartman, 1964)
Potentially Misidentified Species
Ecology
General:
Epigamia noroi, like other members of the Syllidae, reproduces by epitoky. During epitoky the worm undergoes stolonization, in which segments of the body become modified for carrying gametes and swimming (modified setae, enlarged eyes). Sexes are separate and the stolons break off from the stem portion of the worm's body and swim in the water column, as epitokes, with new heads, three antennae, and well-developed eyes. Then, stolons are regenerated. Male and female stolons are morphologically distinguishable (Imajima 1966b; Nygren 2004). The details of fertilization are not known. It is not clear whether the female carries the fertilized eggs in the body or in a ventral egg sac (Imajima 1966b). Details of development are not known, but in the related genus Autolytus, the larvae are brooded and are lecithotrophic (Pettibone 1963).
Epigamia noroi occurs in cold-temperate to warm-temperate climates, from the intertidal to 40 m. It is known from seaweed colonies, intertidal Surfgrass (Phyllospadix spp.) beds, and dock fouling communities (Imajima 1966b; Nygren 2004). Syllids have been considered exclusively carnivorous, including members of the subfamily Autolytinae (inlcuding Epigamia) feeding largely on hydroids (Fauchald and Jumars 1979). However, studies of fecal pellet of Syllids indicate that many, including one species of Autolytus are omnivorous (Giangrande et al. 2000).
Trophic Status:
Omnivore
OmniHabitats
| General Habitat | Rocky | None |
| General Habitat | Marinas & Docks | None |
| Salinity Range | Polyhaline | 18-30 PSU |
| Salinity Range | Euhaline | 30-40 PSU |
| Tidal Range | Low Intertidal | None |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Epibenthic | None |
| Vertical Habitat | Planktonic | None |
Tolerances and Life History Parameters
| Maximum Length (mm) | 13 | Imajima 1966b |
| Broad Temperature Range | None | Cold temperate-Warm temperate |
| Broad Salinity Range | None | Polyhaline-Euhaline |
General Impacts
No impacts have been reported from introduced populations.Regional Distribution Map
Non-native
Native
Cryptogenic
Failed
Occurrence Map
References
Blake, James A.; Ruff, R. Eugene (2007) The Light and Smith Manual: Intertidal invertebrates from Central California to Oregon (4th edition), University of California, Berkeley CA. Pp. 309-410Fauchald, Kristian; Jumars, Peter A. (1979) The diet of worms : A study of polychaete feeding guilds, Oceanography and Marine Biology, an Annual Review 17: 193-284
Giangrande, Adriana; Licciano, Margheritia; Pagliaria, Patrizia (2000) The diversity of diets in Syllidae (Annelida: Polychaeta), Cahiers de Biologie Marine 41: 55-65
Imajima, Minoru (1966b) The Syllidae (polychaetous annelids) from Japan II. Autolytinae, Publications of the Seto Marine Biological Laboratory 14(1): 27-83
Nygren, Arne (2004) Revision of Autolytinae (Syllidae: Polychaeta)., Zootaxa 680: 1-314
Pettibone, Marian H. (1963) Marine polychaete worms of the New England region. 1. Aphroditidae through Trochochaetidae., Bulletin of the United States National Museum 227(Part 1.): 1-356
San Martin, Guillermo; Aguado, Maria Teresa; Alvarez-Campos, Patricia (2014) Revision of the genus Megasyllis (Annelida, Syllidae) with descriptions of four new species, Journal of the Marine Biological Association 94(2): 331-351