Invasion History
First Non-native North American Tidal Record: 1993First Non-native West Coast Tidal Record: 1993
First Non-native East/Gulf Coast Tidal Record:
General Invasion History:
The carnivorous calanoid copepod Tortanus dextrilobatus is native to the coast of China from the Taiwan Strait to the mouth of the Yellow Sea, and to the south coast of South Korea, but is absent from the Yellow Sea itself. This species is characteristic of low-salinity coastal waters (Ohtsuka et al. 1992; Ohtsuka and Reid 1998; Sakaguchi et al. 2011).
North American Invasion History:
Invasion History on the West Coast:
In 1993, this copepod was discovered in western Suisun Bay, California at 3.6 PSU, and by 1999, ranged into San Pablo and the South and Central Bays, over a salinity range of 1.2-28 PSU, reaching densities above 1,000 m-3, which is high for a predatory zooplankter (Orsi and Ohtsuka 1999; Hooff and Bollens 2004). In the lower bay, it was more abundant in San Pablo Bay and South San Francisco Bay than in the central bay (Bollens et al. 2011). It has not been reported from other West Coast estuaries. It has been found in ballast water of ships entering Puget Sound, Washington, but only in those coming from other West Coast ports, presumably San Francisco Bay (Lawrence and Cordell 2010).
Description
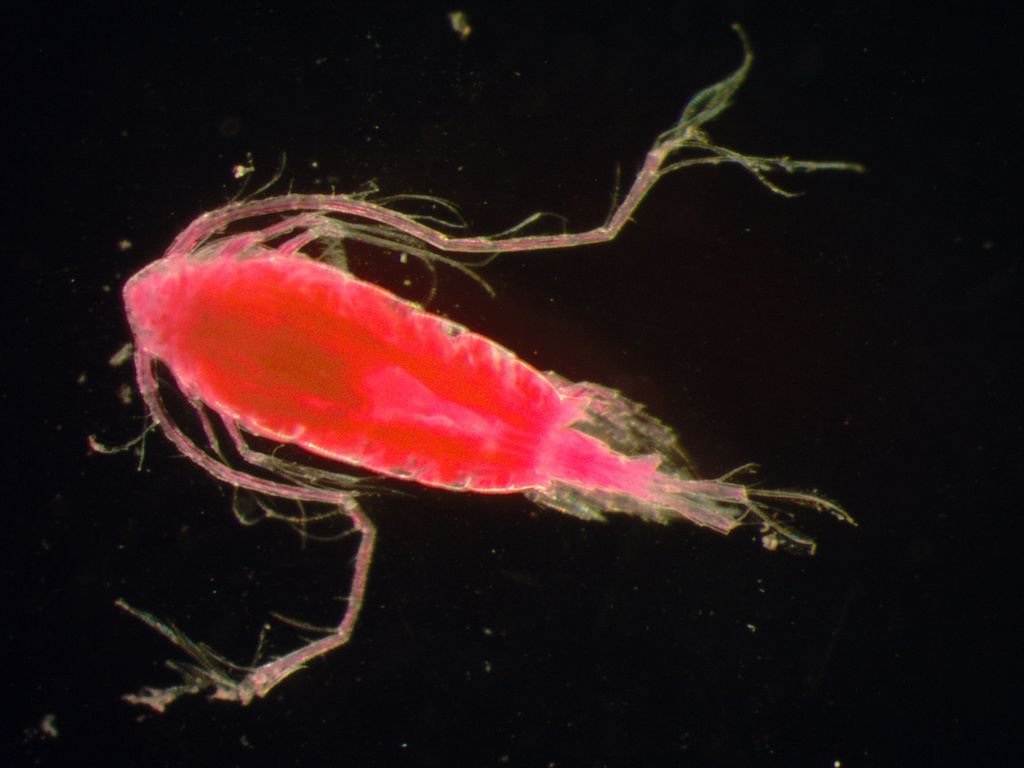
Both sexes of Tortanus dextrilobatus have some asymmetry in the urosome and caudal rami. The feeding appendages are claw-like and prominent, indicative of predatory feeding. Adult females have an oblong cephalothorax, with a somewhat triangular forehead. The head is not fused with the 1st thoracic segment, but the 4th and 5th thoracic segments are fused. There is a single median eye, which is red. The corners of the last thoracic segment form curved, symmetrical protuberances. The urosome has 3 segments. The genital segment is about as long as it is wide, with the genital operculum located ventromedially. The operculum is semicircular, with the posterior margin slightly concave. The 2nd urosomal segment has a posterior-dorsal process on the right side, while the 3rd segment has a larger thorn-like dorso-lateral process. The right caudal ramus is slightly longer than the left one. The two antennules are symmetrical, have 17 segments, and when folded back, reach to the 3rd urosome segment. The exopod and endopod of the 5th leg (P5) consist of only one segment each. The tip of the endopod is bifurcated, and the exopod is claw-like. The length of Chinese specimens was 2.00-2.25 mm (Chen and Zhang 1965), while Korean specimens measured 1.75-1.79 mm, and San Francisco Bay females were 1.58-1.61 mm in size (Ohtsuka et al. 1992; Ohtsuka and Reid 1998; Hoof and Bollens 2004).
Adult males have a much more slender body than the female's. The 4th and 5th thoracic segments are partially fused, and the corners of the 1st thoracic segment are rounded, without protuberances. There are 5 urosome segments, but the posterior segment is very short. As in the female, the right caudal ramus is slightly longer than the left. The right 1st antenna is geniculate (hinged) between the 14th and 15th and comprises 16 segments, but two of these are partially fused. The 5th legs (P5) are each composed of 3 segments, and are asymmetrical. The left leg has broad 1st and 2nd segments, and the distal end of the 3rd segment is curved inwardly. It has setae on the inner surface and 9-10 transverse ridges on the outer surface. The right P5 leg is clawlike. The 1st segment is very broad, with a blunt triangular process on the inner border and a spine at the apex. The distal segment is curved, with 5 denticles and 19-21 fine transverse ridges on the inner surface. Males in China were 2.00-2.25 mm in size (Chen and Zhang 1965) and 1.53-1.76 mm in Korea (Chen and Zhang 1965; Ohtsuka et al. 1992).
The copepodite and naupliar stages of this copepod have not been described, but probably resemble those of Tortanus discaudatus (Johnson 1934). Tortanus dextrilobatus is characteristic of brackish waters (Ohtsuka et al. 1992).
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Arthropoda | |
| Subphylum: | Crustacea | |
| Class: | Maxillopoda | |
| Subclass: | Copepoda | |
| Order: | Calanoida | |
| Family: | Tortanidae | |
| Genus: | Tortanus | |
| Species: | dextrilobatus |
Synonyms
Potentially Misidentified Species
Native to the NW Atlantic and N Pacific, found in poly-euhaline waters
Tortanus forcipatus
Native to NW Pacific, found in poly-euhaline waters
Ecology
General:
Planktonic calanoid copepods mate in the water column. Males use their modified antenules and 5th pair of swimming legs to grasp the female and transfer spermatophores to the female's genital segment. Female copepods of the genus Tortanus lay eggs singly in the water column (Barnes 1983; Cordell et al. 1992; Cordell et al. 2007). Eggs hatch into nauplii which go through six stages. The first stage, NI, has 3 pairs of appendages and is unsegmented - each molt has additional appendages and/or more differentiation of segments. The sixth stage (NVI) molts into a first copepodite stage (CI), with the basic form of the adult and fully differentiated feeding structures, but with only two pairs of swimming legs and only one urosomal segment. The copepod goes through five additonal molts, with increasing numbers of swimming legs, urosomal segments, and sexual differentiation. The sixth (CVI) stage is the male or female adult (Johnson 1934; Barnes 1983).
Tortanus dextrilobatus is characteristic of brackish waters (Ohtsuka et al. 1992; Sakaguchi et al. 2011). Adult T. dextrilobatus feed on smaller copepods, including the native calanoid Acartia spp. and the introduced cyclopoid Oithona davisae. Predation rates increase with temperature and prey density (Hoof and Bollens 2004).
Food:
Copepods
Trophic Status:
Carnivore
CarnHabitats
| General Habitat | Unstructured Bottom | None |
| Salinity Range | Oligohaline | 0.5-5 PSU |
| Salinity Range | Mesohaline | 5-18 PSU |
| Salinity Range | Polyhaline | 18-30 PSU |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Planktonic | None |
Tolerances and Life History Parameters
| Minimum Temperature (ºC) | 8 | Field Data, San Francisco Bay CA (Hooff and Bollens 2004) |
| Maximum Temperature (ºC) | 22 | Field Data, San Francisco Bay CA (Hooff and Bollens 2004) |
| Minimum Salinity (‰) | 0.5 | Field Data, San Francisco Bay CA (Orsi and Ohtsuka 1999) |
| Maximum Salinity (‰) | 28 | Field Data, San Francisco Bay CA (Hooff and Bollens 2004) |
| Maximum Length (mm) | 2.2 | Adult females, 2.05- Adult males(Chen and Zhang 1965); |
| Broad Temperature Range | None | Warm temperate |
| Broad Salinity Range | None | Oligohaline-Polyhaline |
General Impacts
Ecological ImpactsThe predatory copepod Tortanus dextrilobatus has had a significant predatory impact on zooplankton in the San Francisco estuary. Predatory impacts of T. dextrilobatus on native Acartia spp. averaged a 3% reduction of individuals per day for 1997-1999, peaking at 63%. Tortanus dextrilobatus predatory impact on the introduced Oithona davisae averaged 1% per day, peaking at 22% (Hooff and Bollens 2004). The extent to which the invasion of T. dextrilobatus has reduced overall copepod abundance, or higher trophic levels (e.g. fishes), is not clear, given the many other changes occurring in the estuary's pelagic food web.
Regional Impacts
| NEP-V | Northern California to Mid Channel Islands | Ecological Impact | Predation | ||
| Predatory impact of T. dextrilobatus on native Acartia spp. averaged a decrease of 3% of individuals per day for 1997-1999, peaking at 63%; and on introduced Oithona davisae averaged 1% decrease per day, peaking at 22% (Hooff and Bollens 2004). | |||||
| P090 | San Francisco Bay | Ecological Impact | Predation | ||
| Predatory impact of T. dextrilobatus on native Acartia spp. averaged a decrease of 3% of individuals per day for 1997-1999, peaking at 63%; and on introduced Oithona davisae averaged 1% decrease per day, peaking at 22% (Hooff and Bollens 2004). | |||||
| CA | California | Ecological Impact | Predation | ||
| Predatory impact of T. dextrilobatus on native Acartia spp. averaged a decrease of 3% of individuals per day for 1997-1999, peaking at 63%; and on introduced Oithona davisae averaged 1% decrease per day, peaking at 22% (Hooff and Bollens 2004)., Predatory impact of T. dextrilobatus on native Acartia spp. averaged a decrease of 3% of individuals per day for 1997-1999, peaking at 63%; and on introduced Oithona davisae averaged 1% decrease per day, peaking at 22% (Hooff and Bollens 2004). | |||||
Regional Distribution Map
Non-native
Native
Cryptogenic
Failed
Occurrence Map
References
Bollens, Stephen M.; Breckenridge, Joanne K.; Cordell, Jeffery R. Simenstad, Charles A.; Kalata, Olga (2014) Zooplankton of tidal marsh channels in relation to environmental variables in the upper San Francisco Estuary, Aquatic Biology 21: 205-219Bollens, Stephen M.; Breckenridge, Joanne K. Vanden Hoof, Rian C.; Cordell, Jeffery R. (2011) Mesozooplankton of the lower San Francisco Estuary: spatio-temporal patterns, ENSO effects and the prevalence of non-indigenous species, Journal of Plankton Research 33(9): 1358-1377
Bollens, Stephen M.; Cordell, Jeffrey R.; Avent, Sean: Hooff, Rian (2002) Zooplankton invasions: a brief review, plus two case studies from the northeast Pacific Ocean., Hydrobiologia 480: 87-110
Chen, Q.; Zhang, S. (1965) The planktonic copepods of the Yellow Sea and East China Sea I. Calanoida, Studia Marina Sinica 7: 20-31
Cohen, Andrew N.; Carlton, James T. (1995) Nonindigenous aquatic species in a United States estuary: a case study of the biological invasions of the San Francisco Bay and Delta, U.S. Fish and Wildlife Service and National Sea Grant College Program (Connecticut Sea Grant), Washington DC, Silver Spring MD.. Pp. <missing location>
Hooff, Rian C.; Bollens, Stephen M. (2004) Functional response and potential predatory impact of Tortanus dextrilobatus, a carnivorous copepod recently introduced to the San Francisco estuary., Marine Ecological Progress Series 277: 167-179
Huang, Zongguo (Ed.), Junda Lin (Translator) (2001) Marine Species and Their Distributions in China's Seas, Krieger, Malabar, FL. Pp. <missing location>
Johnson, Martin W. (1934) The life history of the copepod Tortanus discaudatus, Biological Bulletin 67(1): 182-200
Kimmerer, William J.; Burau, J. R.; Bennett, W. A. (1998) Tidally oriented vertical migration and position maintenance of zooplankton in a temperate estuary., Limnology and Oceanography 43(7): 1697-1709
Lawrence, David J.; Cordell, Jeffery R. (2010) Relative contributions of domestic and foreign sourced ballast water to propagule pressure in Puget Sound, Washington, USA, Biological Conservation 143: 700–709
Ohtsuka, Susumu; Reid, Janet, W. (1998) Phylogeny and zoogeography of the planktonic copepod genus Tortanus (Calanoida: Tortanidae), with establishment of a new subgenus and descriptions of two new species., Journal of Crustacean Biology 18(4): 774-807
Ohtsuka, Susumu; Yoon, Yang Ho; Endo, Yukihiro (1992) Taxonomy on brackish copepods in Korean waters. 1. Redescription of Tortanus dextrilobatus Chen and Zhang 1965 from Korean waters with remarks on the zoogeography of the subgenus Eutortanus, Journal of the Oceanolgical Society of Korea 27(2): 112-122
Orsi, James J., Ohtsuka, Susumu (1999) Introduction of the Asian copepods Acartiella sinensis, Tortanus dextrilobatus (Copepoda:Calanoida), and Limnoithona tetraspina (Copepoda: Cyclopoida) to the San Francisco Estuary,California, USA., Plankton Biology and Ecology 46(2): 128-131
Rilov, Gil (2009) Biological Invasions in Marine Ecosystems., Springer-Verlag, Berlin Heidelberg. Pp. 261-285
Ruiz, Gregory M.; Geller, Jonathan (2018) Spatial and temporal analysis of marine invasions in California, Part II: Humboldt Bay, Marina del Re, Port Hueneme, and San Francisco Bay, Smithsonian Environmental Research Center & Moss Landing Laboratories, Edgewater MD, Moss Landing CA. Pp. <missing location>
Sakaguchii, Sakiko Orui; Ueda, Hiroshi; Ohtsuka, Susumu; Soh, Young Ho; Yoon, Ho Yoon (2011) Zoogeography of planktonic brackish-water calanoid copepods in western Japan with comparison with neighboring Korean fauna, Plankton & Benthos Research 6: 18-25
Winder; Monika; Jassby, Alan D.; Mac Nally, Ralph (2011) Synergies between climate anomalies and hydrological modifications facilitate estuarine biotic invasions, Ecology Letters 14: 749-757